| 前頁 | 目次 | 次頁 |
|
環境庁国立公害研究所 放射性同位兀素利用による汚染環境の評価と改善に関する研究
〔まえがき〕
環境汚染の傾向を早期に感知し、大事に至る以前にその対策を講じるためには、極低濃度の汚染物質の生物への影響を究明し、汚染環境の評価法を確立する必要がある。また低濃度の汚染物質の生体内での挙動あるいは自然生態系での移行を把握しなければならない。 これらの目的のためにラジオアイソトープをトレーサーとして利用し、生物への微小な影響の検知、及び障害発現の機作の解明に関して研究を行い、汚染環境の評価と改善の手法の開発を検討している。本稿ではこれまでに得られた研究成果のうち、表題にかかげる内容について述べ、研究の現状と今後の予定についても触れる。 〔研究の成果〕
大気汚染環境の植物影響の解析
(1) 気孔閉口反応におけるアブサイシン酸の役割について
植物が大気汚染物質により影響を受けるとき、数段階の防御機構で抵抗する。植物と大気環境との接点である気孔は、大気汚染物質の侵入を防ぐ第一の防壁である。気孔開閉制御ホルモンであるアブサイシン酸(ABA)を多く含む植物が、二酸化硫黄(SO2)や二酸化窒素(NO2)に対して気孔を閉ざして侵入を防ぎ抵抗性を示すことが明らかにされているが、その時のABAの作用機作は不明である。そこで14C-ABAをトレーサーとして使用し、その機作の解明を行った。その結果を、図1及び図2に示す。図1には種々のpHにおける気孔開度に及ぼすABAの影響を示してある。図2には14C-ABAの孔辺細胞への取り込みを種々のpHで示してある。気孔はpH4で最も閉じており、その時ABAも最も多く取り込まれている。気孔閉口とABA取り込み量は平行している。この結果からNO2やSO2が植物体内に侵入し、細胞液に溶けて亜硫酸イオンや亜硝酸イオンになり、pHを低下させると、葉肉細胞から孔辺細胞にABAが移動し、気孔閉口が起こることが示唆された。次に孔辺細胞のABA濃度が高まれば、何故気孔が閉じるかを探るために、14C-グルコースを用いて、孔辺細胞及び孔辺細胞細胞壁の糖代謝活性へのABAの影響を調べた。その結果、ABAが孔辺細胞の糖代謝系及び細胞壁の合成系を阻害し、気孔の閉鎖を引き起こしている可能性が示唆された。 図1. 気孔開度に及ぼすABAの影響 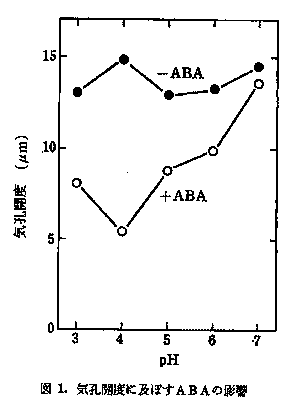 図2. 孔辺細胞へのABAの取り込み 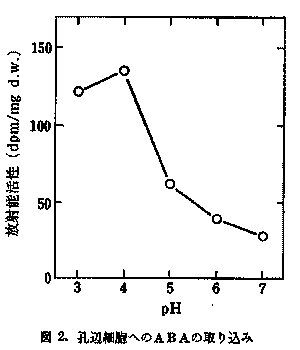 (2) 光合成電子伝達系への影響
植物が大気汚染物質に接触すると、葉に可視障害が現れるが、それ以前にすでに生理機能への障害が起こっている。植物の光合成機能についても阻害が起こるが、その阻害が電子伝達反応なのか、これと共役している光リン酸化反応なのか、あるいは炭酸固定反応なのかについては、区別して明らかにされていなかった。この点を解明するために、32Pを使用して光リン酸化反応への影響を形成されるAT32Pを測定して調べた。結果を表1に示す。 表1 non-cyclicおよびcyclicな光リン酸化反応におよぼすSO2の影響  光リン酸化反応には、水からNADPへ至る、光化学系Ⅰと光化学系Ⅱを直列につないだ電子の流れと共役して起こるnon-cyclicな光リン酸化と、光化学系Ⅰだけで起こる、例えばフェナジンメトサルフェイト(PMS)をcofactorとしたような、cyclicな電子の流れと共役したcyclicな光リン酸化があるが、両者に対するSO2の影響をみたものである。水からNADPまでの電子伝達反応が約36%阻害されている場合に、これと共役するnon-cyclicな光リン酸化も38%阻害を受けている。ところが、光化学系ⅠとⅡの間の電子の流れをジクロロフェニールジメチルウレア(DCMU)で止めて、更にPMSを加えた光化学系Ⅰだけでのcyclicな光リン酸化反応は全く阻害されていない。このことはSO2により阻害を受けているのは電子伝達反応であり、その阻害部位は光化学系Ⅱの周辺にあることを意味する。 次に、SO2の系Ⅱの阻害の機作を明らかにするために、35Sで標識した亜硫酸イオンの葉緑体チラコイド膜への取り込みを、明暗及び明+DCMUの3つの条件で調べた。表2にみられるように、系Ⅱの阻害のかかる明条件下で35Sの取り込みが最大で、DCMUで電子伝達が阻害されると取り込みが半分以下に抑えられ、暗黒下では明条件下の約1/4であった。これらのことは、SO2から生じる亜硫酸イオンが明条件下で多く取り込まれ、これがチラコイド膜のタンパク質部分に直接結合して、阻害を引き起こしている可能性を示唆している。この可能性を支持するデータが、クロロフィル蛍光の活導期現象の解折からも得られている。 表2 35Sのチラコイド膜への取り込み  大気汚染環境の動物影響の解析
(1) 赤血球成分の代謝に及ぼす影響
二酸化窒素(NO2)の生体影響を解明するために、実験動物を用いて研究を行っているが、これまでの研究で、NO2に暴露されたラットの赤血球成分が多様な変化を受けることがわかっている。吸入されたNO2は血液中に硝酸イオンと亜硝酸イオンの形で溶解し、血液中の硝酸イオンレベルはNO2暴露を続ける限り上昇していることが数種の実験動物について観察されている。そこで硝酸イオンの赤血球に対する作用機作を解明するために、赤血球膜リン脂質の脂肪酸代謝に及ぼす影響を、14C-アラキドン酸と3H-パルミチン酸を用いて調べた。その結果、0.1mM以上の硝酸イオンが存在すれば赤血球膜リン脂質中のアラキドン酸含量が増加し、この増加は、硝酸イオンが赤血球へのCa2+イオンの流入を促進し、脂肪酸の取り込みを活性化するためと考える。 一方、NO2暴露により赤血球の血流中での老化が促進されることが明らかになっているが、この老化に膜の脂質代謝が関与しているかどうかを3H又は14C標識脂肪酸を用いて解析した。その結果、赤血球老化に伴い膜への脂肪酸取り込み活性が低下し、膜リン脂質脂肪酸組成が変化することが示唆された。 (2) 肝ミクロソーム成分に及ぼす影響
血液中に溶解したNO2から由来する硝酸イオンは肝臓のミクロソームに局在している薬物代謝系の酵素活性を特異的に減少させることが見出された。この作用機作を明らかにするために、放射性アミノ酸で標識したミクロソームを硝酸イオンとインキュベートして細分画し、放射能の分布を調べた。 ラットに3H-ロイシンを静注して2日間飼育した後14C-ロイシンを静注すると、3Hで標識されたミクロソームは合成されてから時間が経過した蛋白質の多いミクロソームであり、14Cで標識されたミクロソームは合成された直後の蛋白質と考えられる。ミクロソームを細分画した結果、ミクロソームの比重の小さい画分は14Cを含む割合は少なく時間の経過した蛋白質を多く含むものであることがわかった。表3に3H-及び14C-ロイシンで標識したミクロソームを硝酸ナトリウムとインキュベーションすることにより溶出した放射能を示した。添加する硝酸ナトリウムの濃度に依存して3Hで標識した蛋白質の溶出量が増加した。一方、14C標識蛋白質の溶出は濃度によって影響を受けなかった。以上の結果から、NO2暴露によって血液中の硝酸イオン濃度が上昇し肝臓内にも反映されれば、ミクロソーム中の古い蛋白質が溶出し、これが電子伝達系酵素活性の減少を招いていることが明らかにされた。蛋白質の溶出は、硝酸イオンのカオトロピック作用ではないかと推察している。 表3 硝酸ナトリウム処理によるミクロソーム蛋白質の溶出  〔研究の現状〕
当研究所では、上述の研究課題の他に、水生植物による無機塩類及び重金属類の吸収と濃縮に関する研究、水界生態系における汚染物質の動態に関する研究も行っている。重金属、特に水銀を特異的に蓄積するコケ植物(チャツボミゴケ、ムラサキヒシャクゴケ)を発見し、その濃縮特性と蓄積する水銀の存在状態を解析している。これらのコケは生体中に乾重量当り最高1%の濃度まで濃縮蓄積するので、このコケを利用して、物理化学的測定でも検出不可能な、極低濃度の水中の水銀汚染も、生体に濃縮させてモニターすることができるものと考えられる。また、生体中に高濃度に蓄積された水銀が、生理機能に障害を与えていない様子なので、その無毒化された存在状態も興味深い。現在その化学形態はHg-S化合物であることが明らかになったが、その形成機構は現在解明中である。一方、ビーカーレベルで、細菌-クロレラ-ミジンコ-ユスリカから成るモデル生態系を用いて重金属や化学物質の移行、蓄積、循環等について、食物連鎖に注目して研究を進めている。 〔今後の予定〕
大気汚染物質の生物影響の機作に関しては、かなりの点が明らかになってきたが、今後は大気汚染ガスと粉塵中の重金属の複合影響、あるいは光質の変化(オゾン層破壊による紫外線量の増加等)による動植物への影響等を加えて、その阻害機構、無害化機構、障害修復機構、適応、抵抗性獲得の機構について明らかにして行く。 |
| 前頁 | 目次 | 次頁 |